INTRODUCTION
The introduction of nasal endoscopy several decades ago has advanced the diagnosis and treatment of chronic rhinosinusitis (CRS). However, the pathogenesis has remained elusive. Recently, it was proposed that microbial persistence, superantigen (SA) production, and host T-cell response may be involved in the development of chronic eosinophilic respiratory mucosal inflammation (1). SA is a bacterial toxin that binds the major histocompatability complex receptor of the antigen presenting cell and T cell receptor. As a result, antigen processing and presentation are bypassed, causing extensive SA-dependent T cell activation, which increases the severity and chronicity of the inflammation (2). In support of the SA hypothesis, specific IgE to staphylococcal enterotoxins are detected in about 50% of nasal polyp (NP) homogenates (3), and staphylococcal exotoxins can be demonstrated in polyps and mucin specimens of CRS with NP via the enzyme-linked immunosorbant assay (ELISA) (4).
Animal models are an essential tool to validate or reject the SA hypothesis. In the dermatitis model, a single intracutaneous injection of staphylococcal enterotoxin B (SEB) elicits a strong inflammatory reaction in the skin and associated lymphoid tissue (5). In interstitial pneumonia models, a single intratracheal instillation of SEB induces interstitial pneumonia in autoimmune mice (6, 7). However, no SA-induced rhinosinusitis model has yet been been reported. According to the SA hypothesis, a single intranasal application of SEB may induce rhinosinusitis characterized by SA-induced inflammation in terms of severity, chronicity, and cellularity.
The aim of the present study was to develop a rat model of rhinosinusitis induced by SEB. SEB was applied to the nose of rats and histologic evidence of SA-induced nasal inflammation was sought.
MATERIALS AND METHODS
Animal Preparation
Fifty-two 4-week-old Sprague-Dawley rats, with no evidence of upper respiratory tract infections were used. Thirty-six rats received SEB as described below and nine rats were sacrificed 1, 5, 14, and 28 days later. Sixteen control rats received phosphate buffered saline (PBS), and four rats were sacrificed at the same times.
The animals were sedated with intraperitoneally administered ketamine hydrochloride (75 mg/kg) and xylazine hydrochloride (10 mg/kg). Forty microliters of SEB (100 ┬Ąg/mL PBS) (Sigma-Aldrich, St Louis, MO, USA) was dispensed into the nasal cavity by placement of small droplets of solution into each nare. We adopted this amount and concentration of SEB on the basis of a pilot study that optimized SEB concentration in terms of the induction of maximum inflammation without mortality. In the control rats, 40 ┬ĄL of PBS was applied intranasally.
At 1, 5, 14, and 28 days, the animals were painlessly euthanized sacrificed with intraperitoneally administered penobarbital (120 mg/kg). We adopted the time intervals of 1, 5, 14, and 28 days on the basis of previous studies on acute or chronic rhinosinusitis animal models (8-10). The animals were perfused intravascularly with 200 mL of 2% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.2) to fix the tissue. Each head was decapitated and immersed in the fixative overnight. The heads were then stripped of eyes, skin, and muscle, and the mandibles were excised. The specimens were decalcified until soft in 0.25 M ethylenediaminetetraacetic acid in PB. The specimens were excised from a plane posterior to the incisor to a plane anterior to the first molar. The resulting middle segments were washed in PB, saturated overnight at 4Ōäā with 30% sucrose in PB, frozen in liquid nitrogen, and cut into coronal 10 ┬Ąm-thick sections on a cryostat from posterior to anterior.
Histology
The principal area of the histologic analysis and the mucosal sampling areas were selected on the basis of a previous study (9). The principal area of the histologic analysis was the middle segment of the sinonasal cavity in coronal sections including the midline septum, upper nasoturbinate, lower maxilloturbinate, and maxillary sinus. Anatomically similar serial sections were chosen from each animal and were stained with hematoxyline and eosin (H-E).
The appearance of inflammatory cell clusters in the sinonasal air space was observed in the H-E stained sections at 40├Ś magnification. The ratio of the number of sinus spaces occupied by inflammatory cell clusters to the total number of sinus spaces of the histologic sections was determined. The number of inflammatory cells infiltrating the lamina propria was counted at 400├Ś magnification from 10 mucosal sampling areas: lateral sinus wall, upper and lower septa, upper nasoturbinate, and lower maxilloturbinate on both sides. The count was determined for each of three sections per rat.
RESULTS
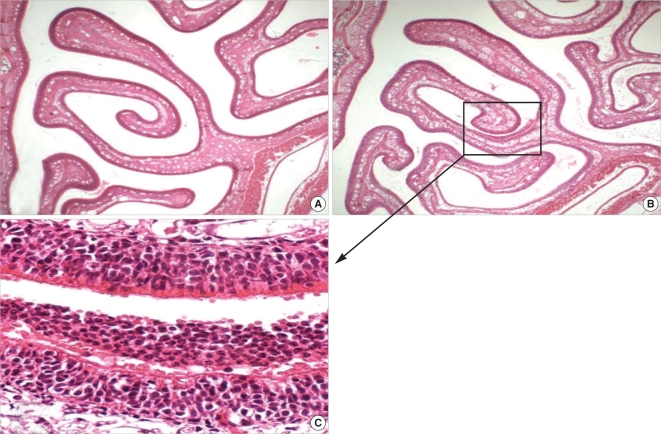
H-E staining revealed no evidence of inflammatory cell clusters in the sinonasal air spaces bounded by the septum, upper naso-turbinate, and lower maxilloturbinate in the sections obtained from control rats (Fig. 1A). In contrast, histologic evidences of rhinosinusitis such as the appearance of neutrophil clusters in the air space and infiltration of neutrophils in the lamina propria were observed in the SEB-applied rats (Fig. 1B, C, and 2B).
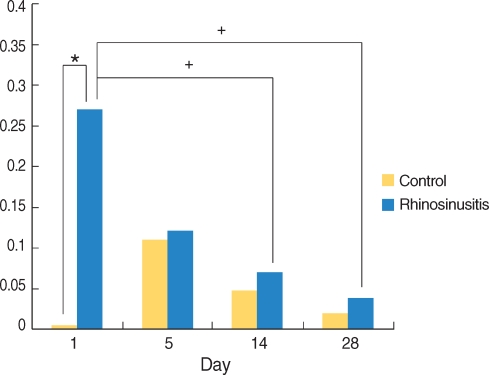
Neutrophil clusters were observed in the sinonasal air space in the SEB-applied rats. The ratio of the sinus spaces occupied by neutrophil clusters was increased significantly at day 1 as compared with the control rats, but regressed by day 5 (Fig. 1B, C, and 3).
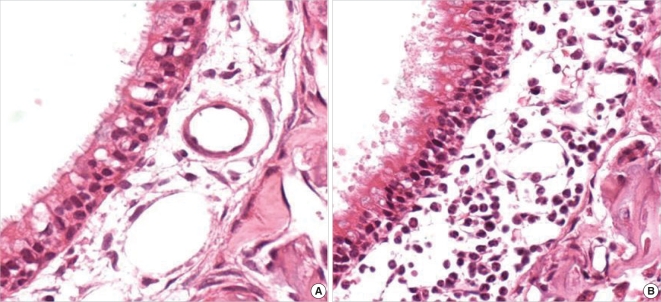
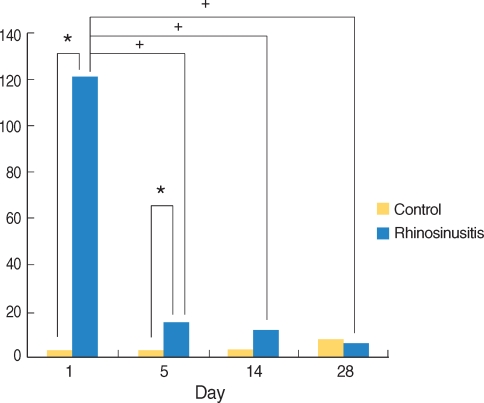
The number of neutrophils infiltrating the lamina propria was increased in the SEB-applied rat. There were significantly more infiltrating neutrophils in the lamina propria of the SEB-applied rats sacrificed at day 1 and 5 compared than the control rats. Comparisons across the full time interval demonstrated statistically significant changes according to the time interval shown by the peak at day 1 (Fig. 2A, B, and 4).
DISCUSSION
In this study, intranasally applied SEB induced acute neutrophilic rhinosinusitis in rats, shown by the peak at day 1, and regressed over five days. Infiltration of monocytes or eosinophils was not observed, nor was there evidence of chronic inflammation lasting more than 14 days. These observations correspond with those from previous studies (11, 12) where, in murine subcutaneous air pouches, SEB can induce an acute inflammation with accumulation of neutrophils predominantly, some monocytes, and a few eosinophils.
Previously, we have reported on the development of rat models for Staphylococcus aureus-induced rhinosinusitis (13) and staphylococcal alpha-toxin-induced rhinosinusitis (14). The histologic characteristics of these models differ from one another. Induction of rhinosinusitis with S. aureus is associated with more severe inflammation with exudate in the sinus air spaces (13), whereas toxin-induced rhinosinusitis is characterized by inflammatory cell clusters, hemorrhage with hemolysis in the sinonasal air spaces, and significant loss of epithelial cells (14). In the present SEB-induced rhinosinusitis rat model, inflammatory cells were predominantly neutrophils, there was no hemorrhage with hemolysis or epithelial damage, and the inflammatory response was maximal at day 1 with a rapid regression thereafter.
Histology of SA-induced inflammation is characterized by infiltration of monocytes and eosinophils, and chronicity. In the dermatitis model, degranulation of dermal mast cells is followed by infiltration of granulocytes including eosinophils, reaching a peak 1 day after SEB injection and regressing over 5-7 days (5). In the interstitial pneumonia model, infiltration of macrophages, monocytes, eosinophils, and fibroblast is observed as early as 3 days after intratracheal administration of SEB, and progresses over the intervening 5 weeks (6, 7). However, in the presently reported SEB-induced rhinosinusitis model, we demonstrate an acute neutrophilic inflammation. We did not observe any histologic evidence of SA-induced inflammation such as infiltration of monocytes and eosinophils, or inflammation lasting more than 14 days. It appears that just the presence of SA in the nose does not necessarily induce SA-induced inflammation, as suggested by the SA hypothesis.
Animal models are essential research tools to understand the pathogenesis of rhinosinusitis. Murine models of rhinosinusitis are well established; most model acute rhinosinusitis (8, 9). In this study, we demonstrate acute neutrophilic inflammation in the nose after the single intranasal application of SEB in rats. A few murine models of chronic rhinosinusitis have been reported. However, they involve surgically manipulation to achieve ostial obstruction (10), or allergy models that require repeated sensitization and challenge (15). There has been no animal model of chronic rhinosinusitis induced by a single causative event capable of pathogenesis as a unified etiology. A very recent study in transgenic mice reported eosinophilic inflammation in the lung after repeated intranasal challenge with low concentration of SEB (16). Further animal studies with transgenic mice or with allergy models induced by repeated intranasal challenge with different concentrations of SEB are needed to prove possible roles of SA in the development or perpetuation of CRS.
In conclusion, we demonstrate that intranasally applied SEB induces acute neutrophilic rhinosinusitis in rats. No evidence of eosinophilic inflammation was apparent. Thus, simple presence of SA in the nose may not be sufficient to induce SA-induced inflammation, in contrast to the SA-hypothesis in dermatitis or pneumonia model.