INTRODUCTION
Age-related hearing loss (ARHL), or presbycusis, is a common problem affecting one-third of adults aged ≥65 years [1]. It is characterized by reduced hearing sensitivity and speech understanding, resulting from degeneration of the cochlea, auditory nerves, or both. ARHL can be affected by genetics, metabolism, noise, ototoxic medications, and inflammation [2]. For example, DCLK1, SLC28A3, CEP104, and PCDH20 are genes associated with hearing function in humans [3]. Reactive oxygen species (ROS) are generated by the production of superoxides and hydrogen peroxide (H2O2), and these have the capability of damaging DNA, leading to a loss of proteins and apoptosis in the cochlea [4]. Exposure to noise and ototoxic drugs can lead to cochlear cell degeneration and hearing loss [5]. Inflammation and inflammatory mediators contribute to an increased risk of hearing impairment [6]. ARHL contributes to social isolation, depression, and dementia in aged adults [7]. Therefore, it is important to explore the mechanism of ARHL and identify new therapeutic targets for clinical treatment to prevent hearing loss and improve quality of life.
While ARHL is considered a multifactorial disorder, many studies have focused mostly on protein-coding messenger RNAs (mRNAs). More recently, thousands of non-coding transcripts have been identified through high-throughput sequencing, and some of them have been shown to be functional. Long non-coding RNAs (lncRNAs) are a type of RNA, generally defined as transcripts of more than 200 nucleotides that are not translated into protein [8]. Many studies reported that lncRNAs can control gene regulation via epigenetic modifications, recruit transcription factors, and interact with splicing factors [9]. Some lncRNAs contain miRNA within their sequences and are referred as miRNA host genes lncRNAs (lnc-miRHGs). These lnc-miRHGs have been reported to serve as competing endogenous RNA (ceRNAs), DNA interactors, and protein interactors. [10]. lncRNAs play key roles in biological processes, and alteration of lncRNAs expression levels is associated with diseases such as cancer [9]. Recent studies have established a direct correlation between lncRNA regulation and ARHL in aged mice. The lncRNA Gm44593 can reduce ROS levels by regulating miR-29b/WNK1 signaling in ARHL [11]. The lncRNA AW112010 regulates mitochondrial function and biogenesis, leading to AMP-activated protein kinase (AMPK) activation in ARHL [12]. Downregulated expression of the lncRNA myocardial infarction associated transcript (MIAT) was found in aged hair cells, and MIAT could regulate the expression of SIRT1/PGC-1a by downregulating miR-29b [13]. In addition, upregulated Sirt1 expression by the lncRNA H19 regulates H2O2-induced cochlear hair cell apoptosis by inhibiting miR-653-5p [14]. Although the functions of a few lncRNAs in ARHL have been revealed, the common lncRNAs involved in the ARHL process remain unknown.
In this study, we investigated common mRNA and lncRNA profiles in ARHL through a bioinformatics analysis. In total, 112 common mRNAs and 10 common lncRNAs were identified as being differentially expressed (DE) in the cochleae of three different RNA-sequencing (RNA-seq) datasets. These findings may help identify effective targets for ARHL.
MATERIALS AND METHODS
Database search and data preprocessing
We searched the GEO database (Gene Expression Omnibus; https://www.ncbi.nlm.nih.gov/geo/) for datasets related to ARHL by using the keywords “aging,” “hearing loss,” and “presbycusis.” Three datasets for the cochlea (GSE127204, GSE196870, and PRJNA600057) were downloaded from European Nucleotide Archive (ENA). FASEQ files were downloaded directly from ENA using the IBM Aspera tool (v4.0.0.182279). FASTQ files were trimmed using Trimmomatic (v0.39) [15] to remove adaptors and low-quality reads. The trimmed reads were aligned and quantified to the Gencode vM26 mouse genome (https://www.gencodegenes.org/) [16] using Salmon (v1.4.0) with default settings [17].
Data preprocessing and differential expression analysis
Quantified transcript abundances using Salmon were converted to gene-level counts using the tximport (v1.6.3) [18] package in R. Differential expression analysis of mRNA and lncRNA between two conditions was performed using the DESeq2 (v1.30.1) [19] package. We selected the top DE genes (DEGs) of the mRNAs and lncRNAs from the mouse cochlea using the P-values calculated using DESeq2 (Supplementary Tables 1 and 2). Each RNA sequencing dataset was analyzed individually.
Visualization of RNA seq data
Venn diagram and heatmap of DE mRNAs and DE lncRNAs were generated using an online tools provided by VIB and Ghent University (https://bioinformatics.psb.ugent.be/webtools/Venn/) and ComplexHeatmap R package (v 2.6.2) [20]. Genome information on the lncRNAs was generated using the Gviz R package (v1.34.1).
Gene Ontology enrichment analysis
To explore the functions of the DEGs in the cochlea of aged mice, Gene Ontology (GO) enrichment analysis was performed using DAVID Bioinformatics Resources 6.8 (https://david.ncifcrf.gov) using 79 upregulated and 33 downregulated genes. GO analysis involved three categories, namely biological processes (BPs), cellular components (CCs) and molecular function (MF) with P-value <0.05.
Construction of the lncRNAs-mRNAs network
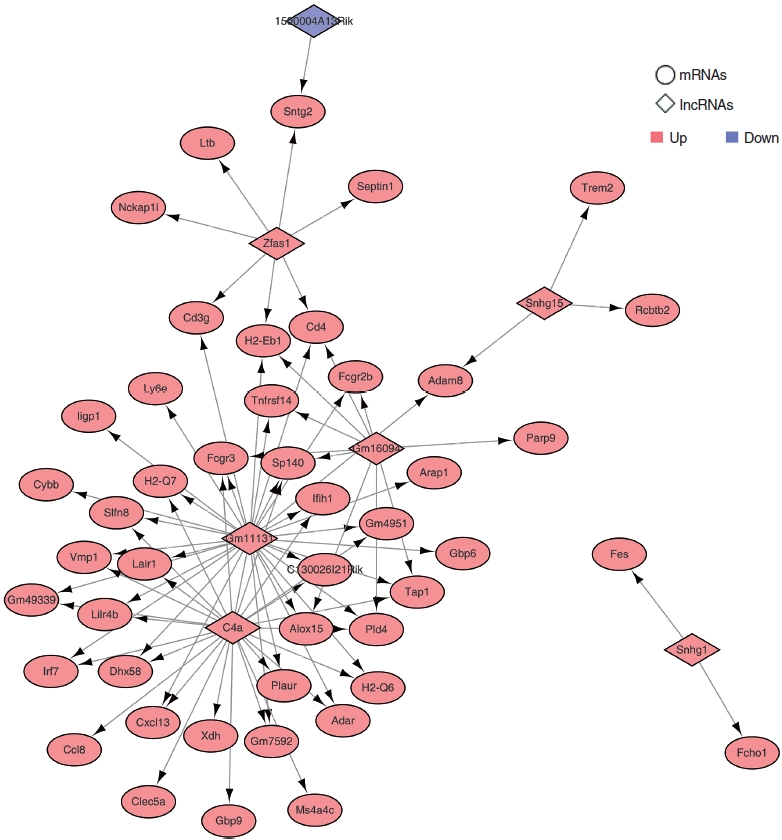
To construct the potential lncRNAs-mRNAs network, we performed Pearson correlation coefficient (PCC) analysis using the Hmisc R package (v4.7.0). PCC was calculated between the 10 lncRNAs and 112 mRNAs with cutoff values of |Pearson correlation| >0.9, P<0.05, and the lncRNA-mRNA regulatory network was structured using Cytoscape 3.8.2. Diamond shape represent lncRNAs and circular (ellipse) shape represent mRNAs. Red nodes represent upregulated lncRNAs or mRNAs, and blue nodes represent downregulated lncRNAs or mRNAs.
HEI-OC1 cell culture
The House Ear Institute-Organ of Corti 1 (HEI-OC1) cells were cultured in high glucose Dulbecco’s Modified Eagle Medium (DMEM; Welgene, #11965092) containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific Inc., #10437028) without antibiotics at 33 °C and 10% CO2.
Validation of mRNAs and lncRNAs by quantitative real-time PCR
To confirm individual mRNA and lncRNA, total RNA was extracted from HEI-OC1 cells treated with 1 mM H2O2 for 6 hours using the RNAiso Plus Reagent (Takara) following the manufacturer’s protocol. cDNA was synthesized using the M-MLV Reverse Transcriptase, RNaseH⁺ premix kit EBT-1511 (Elpisbio) with random hexamers according to the manufacture’s protocol. Real-time PCR was performed with TB Green Premix Ex Taq (Tli RNaseH Plus, Takara Bio Inc.) using a Thermal Cycler Dice Real Time System III (Takara Bio Inc.). PCR analyses were as follows: 95 °C for 30 seconds followed by 40 cycles at 95 °C for 5 seconds and 60 °C for 30 seconds. Relative RNA amounts were calculated using the 2–ΔΔCT method. β-actin (ACTB) was used as an internal control for the relative quantitation of mRNAs. Primer details are listed in in Supplementary Table 3.
Statistical analysis
Data are presented as the mean±standard error of the mean, and statistical analyses were performed using unpaired Student t-test to complete the differential expression analysis of mRNAs and lncRNAs using real-time quantitative reverse-transcription polymerase chain reaction (qRT-PCR). Statistical significance was set at P<0.05.
RESULTS
Identification and functional analysis of mRNAs in ARHL
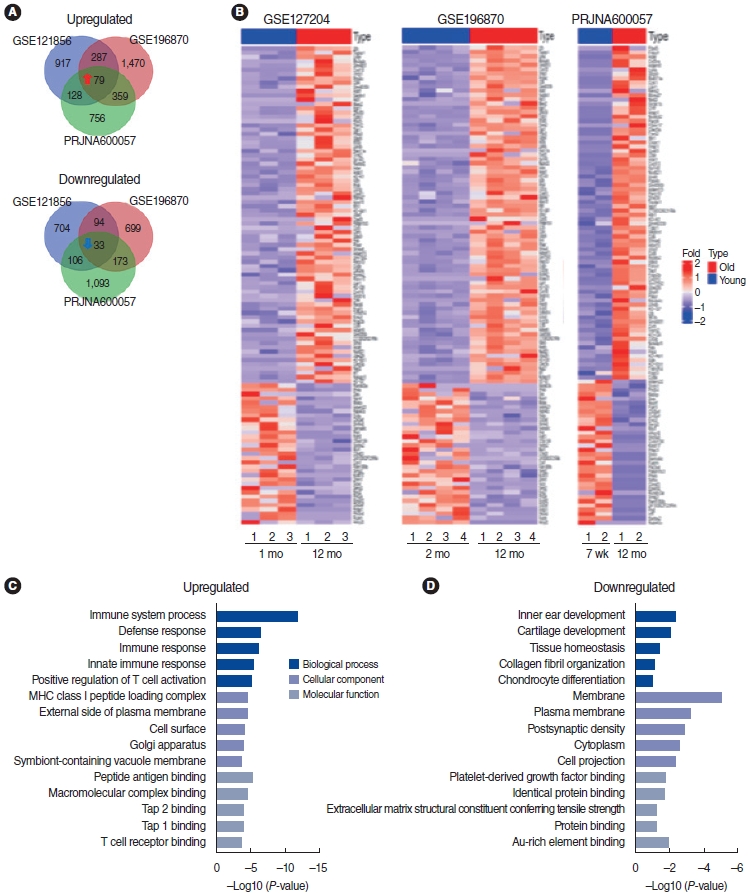
To identify the important role of mRNA gene expression profiles in ARHL, we used publicly available GEO datasets to analyze the differentially expressed genes (DEGs) in the cochleae of aged mice compared to young mice (Table 1). In total, 112 common mRNAs (79 upregulated and 33 downregulated mRNAs) were identified in all three GEO datasets with a cutoff of |log2FC| ≥ 1.5 and P≤0.05. These were visualized using a Venn diagram (Fig. 1A) and heatmap (Fig. 1B). The upregulated and downregulated genes are listed in Supplementary Table 1. Next, we performed GO analysis with the 112 mRNAs to identify the relevant biological function and enriched signaling pathways using the DAVID website. The details of the GO list are presented in Supplementary Table 4. As shown in Fig. 1C and D, the GO analysis of BPs indicated that the 79 upregulated mRNAs participated in immune system processes, the defense response, the immune response, the innate immune response, and positive regulation of T cell activation. The GO analysis of CCs indicated that the genes participated in the major histocompatibility complex (MHC) class I peptide loading complex, the external side of the plasma membrane, the cell surface, the Golgi apparatus, and symbiont-containing vacuole membranes. The GO analysis of MFs indicated that the genes participated in peptide antigen binding, macromolecular complex binding, TAP2 binding, TAP1 binding, and T cell receptor binding (Fig. 1C). The 33 downregulated mRNAs were involved in the following BPs: modulation of inner ear development, cartilage development, tissue homeostasis, collagen fibril organization, and chondrocyte differentiation. In terms of CCs, genes were associated with the membrane, plasma membrane, postsynaptic density, cytoplasm, and cell projection. In terms of MFs, the genes participated in platelet-derived growth factor binding, identical protein binding, extracellular matrix structural constituent conferring tensile strength, and protein binding (Fig. 1D).
Verification of DE mRNA profiling in ARHL
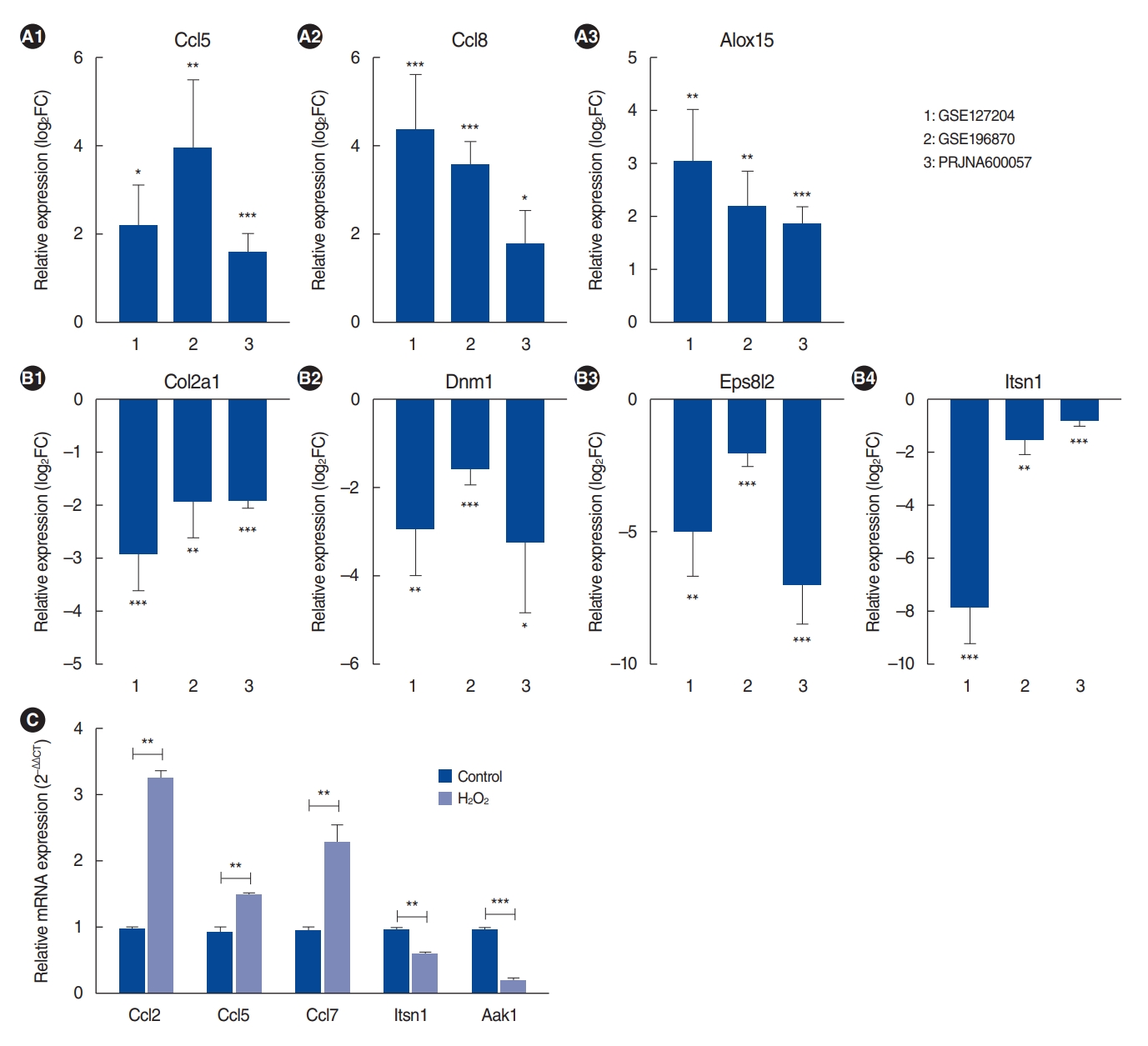
We identified 112 common mRNAs in aged cochleae from the RNA-seq data. Among these encoding defense and immune response genes, Ccl2 (chemokine [C-C motif] ligand 2), Ccl5 (chemokine [C-C motif] ligand 5), and ALOX15 (Arachidonate 15-Lipoxygenase) were upregulated in aged cochleae in the RNA-seq data (Fig. 2A). The inflammation-related gene Alox15 was upregulated in the cochleae of aged C57BL/6 mice [21].
Regarding the regulation of cell communication genes, Col2a1 (collagen type ii alpha 1 chain), Dnm1 (dynamin 1), Eps8l2 (epidermal growth factor receptor kinase substrate 8-like protein 2) and Itsn1 (intersectin 1) were downregulated in aged cochleae in the RNA-seq data (Fig. 2B).
To validate the DE mRNA results, H2O2-treated HEI-OC1 cells were used. H2O2-treated HEI-OC1 cells have been used as an in vitro system to study cellular and molecular mechanisms involved in ARHL [23-26]. We selected three defense responserelated genes and two genes related to the regulation of cell communication and performed qRT-PCR with 1 mM H2O2-treated HEI-OC1 cells. The primer details are presented in Supplementary Table 3. Ccl2, Ccl5, and Ccl7 were significantly upregulated in H2O2-treated HEI-OC1 cells compared with their expression in the control group. Itsn1 and Aak1 were significantly downregulated in H2O2-treated HEI-OC1 cells (Fig. 2C). These qRT-PCR results were consistent with the RNA-seq data. In conclusion, the results showed that the RNA-seq analysis data were reliable, with high specificity for ARHL.
Identification of DE lncRNAs in ARHL
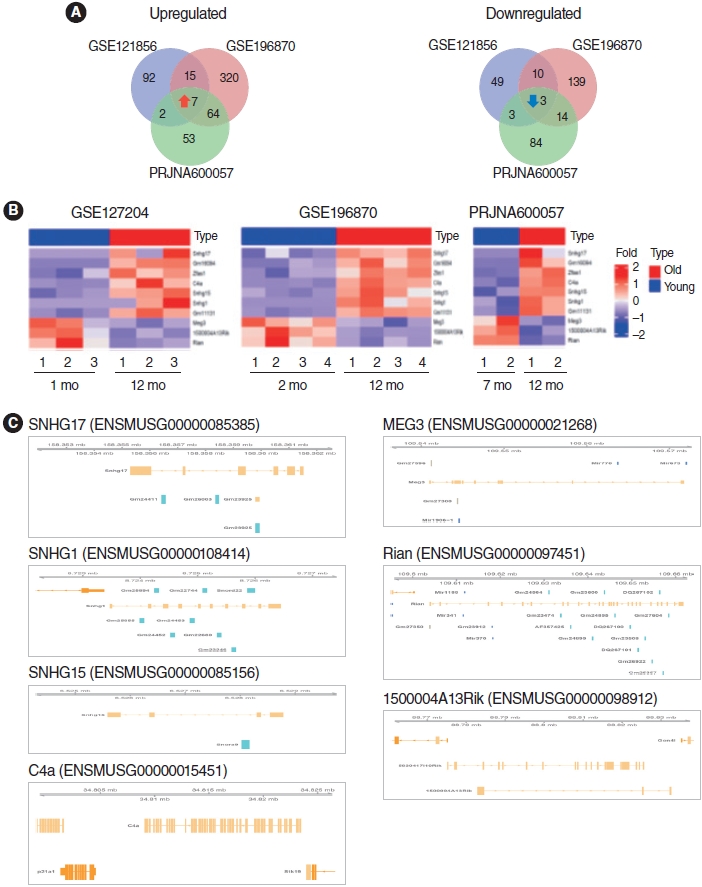
To identify the lncRNAs associated with ARHL, we analyzed three RNA-seq datasets. We identified a total of 10 lncRNAs, including seven upregulated and three downregulated lncRNAs (P<0.05 and |log2FC|>1.0) (Fig. 3A) in the cochlear of aged mice compared to those in young mice. These DE lncRNAs were visualized using Venn diagrams and heatmaps (Fig. 3A and B, Supplementary Table 2). In addition, the genomic information of DE lncRNAs was displayed using the Gviz R package (Fig. 3C).
Some lncRNAs harbor miRNA sequences and act as lnc-miRHGs. Zhang et al. [27] revealed that the extent and specificity of miRNA expression during cochlear aging contributes to the progression of ARHL. Among the identified lncRNAs, Meg3 and Rian were demonstrated to be lnc-miRHGs. Based on genomic information of the identified lncRNAs, MEG3 contains the miR1906-1 and miR770 sequences, and Rian contains the miR1188, miR341, and miR370 sequences. The functions and regulatory mechanisms of these miRNAs in ARHL are unknown. Therefore, these miRNAs require further investigation.
Verification of DE lncRNA profiling in ARHL
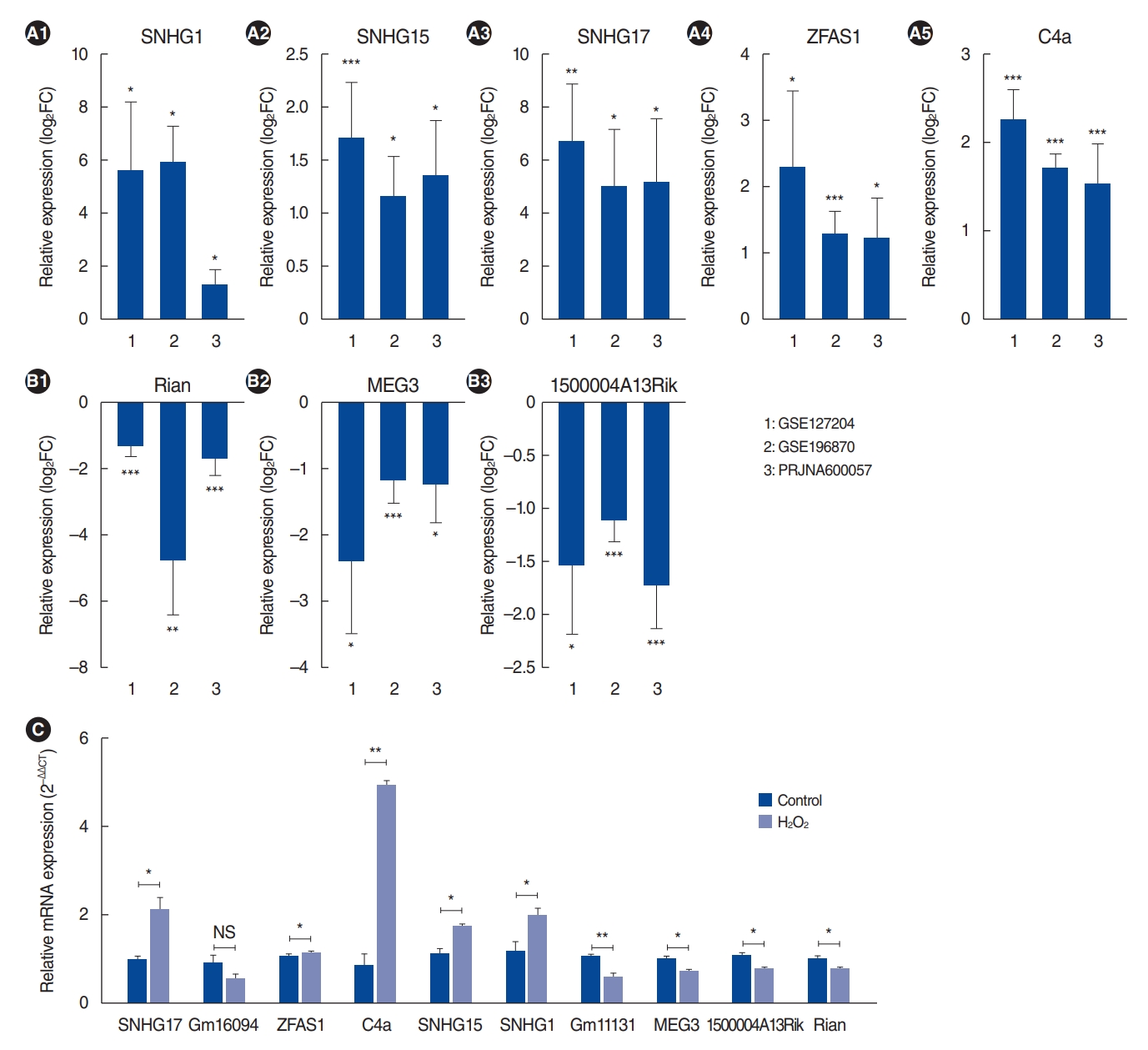
We identified 10 common lncRNAs in aged cochleae in the RNA-seq data. Bar graphs were generated to represent upregulated and downregulated lncRNAs in all RNA-seq datasets (Fig. 4A and B). Among them, Rian has already been reported as an ARHL-associated lncRNA [12]. Consistent with the previous report, Rian expression was significantly downregulated in aged cochleae in our analysis.
To confirm the DE lncRNA results, we performed qRT-PCR validation in H2O2-treated HEI-OC1 cells. As shown in Fig. 4C, the lncRNAs SNHG17, ZFAS1, C4a, SNHG15 and SNHG1 were upregulated in H2O2-treated HEI-OC1 cells, whereas MEG3, 1500004A13Rik, and Rian were downregulated. These qRT-PCR results were all in accordance with the RNA-seq data, except Gm16094 and Gm11131. However, the relationship between these lncRNAs and ARHL regulation remains unknown.
Identification of lncRNA-mRNA network
To demonstrate which lncRNAs are involved in ARHL, we chose 10 lncRNAs and 112 mRNAs to construct the lncRNA-mRNA interaction network. We performed trans pattern analysis of the PCCs (P<0.05 and |Pearson correlation| >0.9), based on the RNA-seq analysis data using the Hmisc package. We identified 80 common co-expressed pairs of upregulated and downregulated lncRNAs in the three different RNA-seq datasets (Fig. 5, Supplementary Table 5). These co-expressed pairs were used to build a lncRNA-mRNA co-expression network using Cytoscape. In the co-expression network, C4a and Gm11131 showed the highest number of interactions.
DISCUSSION
ARHL is a common problem in aging societies worldwide. The primary pathology of ARHL includes the degeneration of hair cells, stria vascularis, and ganglion neurons in the cochlea, as well as the auditory nerve. Hearing loss caused by genetic factors and exposure to oxidative stress, noise, and ototoxic drugs not only affects hearing sensitivity and speech understanding, but also leads to social isolation, depression, and dementia in older adults. Therefore, understanding the mechanism of ARHL is important for its prevention.
In this study, we focused on the common expression profiles of mRNA and lncRNA in ARHL using a bioinformatics approach. From the mRNA profiles of young and aged mice, we identified 112 common mRNAs from three different RNA-seq datasets. Among them, upregulated mRNAs were related to immune system processes, the defense response, the immune response, the innate immune response, and positive regulation of T cell activation (Fig. 1C). These results closely align with those reported by Chen et al. [22], who revealed that a mouse model based on aging, noise exposure, and cisplatin ototoxicity shared hub genes involved in immune system processes, the inflammatory response, and apoptotic processes. Similarly, the 79 upregulated mRNAs identified in this study were involved in defense and immune responses. Increased levels of ROS in aging result in the accumulation of oxidative stress, leading to mitochondrial damage [28]. Oxidative stress is one of the pathologies linked to ARHL, and ROS can modulate immune response and inflammation by inducing oxidative stress [6,29].
We identified 10 common lncRNAs from three different RNA-seq datasets. Among them, Rian had been reported to be downregulated in the cochleae of aged mice. However, the other nine lncRNAs were not known to have any roles in ARHL. Small nucleolar RNA host genes (SNHGs) are a group of long noncoding RNAs, which are reported in many studies as being overexpressed in various cancers. With very few exceptions, the members of the SNHG family are recognized as inducing increased proliferation, cell cycle progression, invasion, and metastasis of cancer cells through interaction with methyltransferase and transcription factors and act as ceRNAs for miRNAs [30]. The lncRNA SNHG17 has been reported to enhance glioma cell proliferation, migration, and invasion by inducing zinc fingers and homeoboxes 1 (ZHX1) expression via inhibiting miR-23b-3p [31]. The lncRNA SNHG1 promotes cervical cancer progression by regulating NIMA-related kinase 2 (NEK2) expression via inhibiting miR-195 [32]. In addition, SNHG1 enhances cell proliferation, migration and invasion in esophageal cancer by regulating cell division cycle 42 (Cdc42) expression via inhibiting miR-195 [33]. lncRNA SNHG15 was increased in human cancers and can contribute to cell proliferation by acting as a sponge for miRNAs [34]. We found that the lncRNAs SNHG1, SNHG15, and SNHG17 were upregulated in both aged cochleae and H2O2-treated HEI-OC1 cells. mir-23b-3p and miR-195, which could act as ceRNAs of SNHG1 and SNHG17, were reported to be downregulated in the cochleae of aged C57BL/6 mice [27]. However, the functional roles of these miRNAs and their target genes in the cochleae are not known. In addition, the biological functions of the miRNA-containing lncRNAs MEG3 and Rian are still unknown (Fig. 3C). We identified a total of 80 trans-regulated target genes for upregulated and downregulated lncRNAs from the aged mouse cochleae (Fig. 5). This is the first step in understanding how lncRNAs regulate ARHL. The predicted target genes were highly enriched in biological processes, including immune system processes, the innate immune response, the defense response and the defense response to viruses. These results indicate that the identified lncRNAs and miRNAs could be associated with ARHL. However, the detailed regulatory mechanism needs to be further explored.
In this paper, we used a bioinformatics approach to identify mRNAs and lncRNAs involved in ARHL from public RNA-seq data generated in different environments with animals from different genetic backgrounds and different ages at the time of testing. Our study could provide a deeper understanding of the function of mRNAs and lncRNAs in ARHL development, as well as potential targets for preventing ARHL.
HIGHLIGHTS
▪ We identified 112 common messenger RNAs (mRNAs) and 10 common long non-coding RNAs (lncRNAs) from aged cochlear samples in three different RNA-sequencing datasets.
▪ Age-related hearing loss (ARHL)-associated mRNAs were highly enriched in biological processes such as the immune system and defense response.
▪ The lncRNAs Gm11131, C4a, and Gm16094 were the ARHL-related lncRNAs identified with the highest frequency from aged cochlear samples.